Ji Hye Kim1,
Min Young Kim1,2 ![]()
For correspondence:- Min Kim Email: jeffmkim@jejunu.ac.kr Tel:+82647543349
Received: 19 February 2016 Accepted: 27 October 2016 Published: 20 December 2016
Citation: Kim JH, Kim MY. Effect of dose and dosing rate on the mutagenesis of nitric oxide in supF shuttle vector. Trop J Pharm Res 2016; 15(12):2587-2593 doi: 10.4314/tjpr.v15i12.8
© 2016 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
Purpose: To determine how the dose and rate of NO• treatment affects mutagenic responses.
Methods: Shuttle vector pSP189 was used to determine the genotoxicity resulting from in vitro exposure to NO• using three delivery methods (reactor and Transwell co-culture systems, and NO• donor sodium nitroprusside), followed by plasmid replication in bacteria MBL50 and human AD293 cells.
Results: When exposed to preformed 100% NO• for 3 h or 1% NO• for 35 h using a reactor system, a cumulative dose of 1260 μM × min reduced AD293 cell viability by 46 and 18% and increased mutation frequencies (MFs) 1.9- and 5.3-fold higher than argon control, respectively. Roughly 5-fold increase in MF of the supF gene of AD293 cells co-cultivated with macrophages stimulated with IFN-γ/LPS was also observed. When AD293 cells were treated by SNP, DNA strand breaks were induced and MFs were increased in a dose-dependent manner.
Conclusion: These results provide important clues to how dose and dosing rate of introducing NO• may contribute to potential genotoxicity resulting from NO• formation in vivo.
Introduction
Nitric oxide (NO•) is a key molecule that has significant effects on many physiologic processes in the body [1]. Low concentrations of NO• plays a role in inflammation, immune response, and neurotransmission in the brain, as well as in the functioning of the cardiovascular system [2]. However, long-term exposure to elevated NO• on host and neighboring cells can damage DNA through various mechanisms such as base deamination, induction of strand break and alkylation by N-nitroso compounds [3]. NO• also reacts rapidly with superoxide anion (O2•¯) to form a strong oxidant and nitrating agent, peroxynitrite (ONOO-), which can initiate reactions characteristic of the hydroxyl radical (•OH), nitronium ion (NO2+), nitrogen dioxide radical (NO2•) [4]. These reactive oxygen species (ROS) produced by inflammatory cells have been proposed to contribute to multi stage carcinogenesis by inducing DNA or tissue damage and mutations [4, 5].
Many studies have been performed to clarify the genotoxic effects of NO• and ROS [6-9]. To gain insight into the underlying molecular events that lead to mutations both in vitro and in vivo model systems have been employed in these studies. Shuttle vector plasmid is a useful tool for delivering foreign or modified DNA or for complementing mutations. For these reasons, the shuttle vector carrying the supF gene has been used by other laboratories as well as our group to investigate the mutational specificity and spectra induced by NO• and ROS.
The pathobiochemistry resulting from overproduction of NO• and ROS is thought to be associated with chronic inflammation and carcinogenesis [10]. NO• and ROS produced by activated inflammatory cells during chronic inflammation can alter functionally important biological molecules, resulting in effects such as mutations in oncogenes and tumor suppressor genes or posttranslational modification of proteins associated with an increased risk for a variety of cancers [10]. A critical feature of the link between any genotoxin and cancer is the underlying chemistry, which in the case of NO• is complicated by the possibility that concentration and flux of NO• affect the product distribution and yields. This is illustrated by our recent observations of different nucleobase damage chemistry depending on the means of NO• delivery [11].
The purpose of the present study was to determine how NO• dose rate of treatment delivery affects mutagenic responses. Two NO• delivery systems (reactor and co-culture systems) and a donor compound generating the precursors of NO• provide useful surrogate means for determining DNA damage and mutagenic responses to low levels of NO• administered continuously over substantial time periods. We therefore used the shuttle vector pSP189, to determine mutations resulting from in vitro exposure to NO• using three delivery methods (reactor and co-culture systems, and sodium nitroprusside), followed by plasmid replication in bacterial and human cells, to determine possible effects of differences in repair and replication in the two host cells.
Methods
Cell cultures and chemicals
Mouse macrophage-like RAW 264.7 and AD293 cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM) supplemented with antibiotics and 10% heat-inactivated fetal calf serum (Cellgro, Corning, NY). These cell lines were maintained at 37 ºC in humidified 5% CO2 atmosphere. Other reagents used were: gases from Air Gas (Edison, NJ); Silastic™ tubing (0.058 in. i.d., 0.077 in. o.d.) from Dow Corning (Midland, MI); total NO• immunoassay kit and recombinant mouse IFN-γ from R&D Systems (Minneapolis, MN); Escherichia coli LPS (serotype 0127:B8) and sodium nitroprusside (SNP) from Sigma (St. Louis, MO).
Plasmid amplification
The pSP189 shuttle vector containing an 8-bp ‘signature sequence’ was a gift from Dr. Michael M. Seidman (NIH, Bethesda, MD). As described previously [6], the pSP189 plasmid was amplified in E. coli AB2463 cells grown at 37 °C in LB media with 50 µg/mL ampicillin (Sigma) for 12-14 h with shaking at 250 rpm and was isolated using a Maxi DNA isolation and purification kit (Qiagen, Valencia, CA).
Transfection of AD293 cells
Twenty four hours prior to transfection, AD293 cells were seeded at a density of 5 × 106 in 100 mm tissue culture plates (Falcon). The cells were transfected with pSP189 plasmid DNA containing supF gene (20-40 µg per 100 mm tissue culture plate) according to the protocol for the MBS mammalian transfection kit supplied by Stratagene (La Jolla, CA). After 24 h, the cells were removed from plates after trypsinization and co-cultured with RAW 264.7 cells.
Exposure of AD293 cells to NO• in a reactor system
After transfection with plasmids as indicated, AD293 cells were exposed to NO• by diffusion through permeable SilasticTM tubing utilizing specially designed reactors, with which NO• dose and dosing rate can be tightly controlled at steady state concentrations as described previously [11]. One day prior to treatment, transfected AD293 cells were seeded at a density of 5 × 106 in 60 mm tissue culture plates to allow the cells to adhere, after which they were exposed to 100% NO• or 1% NO• in a reactor system. The total NO• dose delivered into the medium, expressed in units of μM × min, was controlled by varying the exposure time. Cells exposed to argon gas under the same conditions served as negative controls.
Co-culture of AD293 and IFN-γ/LPS-stimulated RAW 264.7 cells
AD293 transfected with plasmids and RAW 264.7 cells were placed together at a ratio of 1:2 (0.5 × 107 : 1 × 107, respectively) in 10 mL of DMEM, and incubated for 12, 24, 48 and 72 h in humidified atmosphere with 5% CO2 at 37 °C. On day 0, the co-cultured cells were treated with 20 units/mL IFN-γ and 20 ng/mL LPS. All cultures were repeated in triplicate.
Treatment of plasmid with NO• donor SNP
SNP was dissolved in 150 mM sodium phosphate buffer, pH 7.4, just before use and 10 µL aliquots were added to 90 µL plasmid solutions in buffer. Final SNP concentrations ranged from 0-4 mM. Suspensions were incubated at 37 °C for 60, 100, or 180 min in a shaking water bath. At the end of treatment, DNA samples were washed twice with cold TE buffer (pH 7.4) using Amicon Centricon-30 concentrators (Millipore, Billerica, MA). The plasmid DNA was stored in TE buffer on ice until the DNA damage analysis and the transformation into MBL50 cells was carried out within 1-2 h of treatment.
Cell viability assay
Cell viability 24 h after treatment was determined by trypan blue exclusion to determine dose-response and time-dependent effects of NO• on AD293 cells.
Analysis of DNA single-strand breaks
The conversion of the covalently closed circular double-stranded supercoiled DNA to an open circle (relaxed) form was used to investigate DNA strand breakage induced by SNP. An aliquot (12 μL) was loaded onto 1.0% agarose gel and electrophoresis was carried out at 100 V.
Determination of NO• levels
Total NO• [nitrate NO3¯ plus NO2¯] and NO2¯ production were measured with a nitric oxide assay kit (R&D Systems). In addition, 50 μL of culture supernatant or plasmid solution was allowed to react with 100 μL of Griess reagent and incubated at room temperature for 10-30 min. For measurement of total NO• production, NADH and NO3¯ reductase were added before reaction with the Griess reagents. Optical density was measured using a microplate reader at 540 nm. Fresh culture media served as the blank in all experiments. Total NO• and NO2¯ concentrations were calculated from standard curves derived from NO3¯ and NO2¯ standard solutions provided with the kit.
Recovery of plasmid from AD293 cells
After each stimulation time, the plasmid was recovered from trypsinized cells by extraction with Promega Wizard miniprep DNA purification kit. Unreplicated input plasmids were removed by digestion with the restriction endonuclease DpnI. MBL50 cells were transformed with aliquots of the recovered DNA.
DNA transformation into MBL50 cells and selection of mutated supF gene
E. coli MBL50 [F¯ CA7020 lacY1 hsdR hsdM galU galK rpsL thi lacZ(Am) ∆(araBAC-leu)7679 araD¯ araC(Am)] was the host for the selection of forward mutations in the target supF gene [6]. This strain was prepared and used for electroporation as previously described [6]. Aliquots of transformed MBL50 cells were plated onto medium A with 50 µg/mL ampicillin, 20 µg/mL IPTG (Roche), and 10 µg/mL X-gal (Roche), supplemented with 2 g/L L-arabinose (Sigma) for selection of mutants. The remaining suspension was diluted and plated onto LB agar containing only ampicillin for determination of total number of transformants. Transformation efficiency (TE) was expressed as the number of colony forming units (cfu) produced by 1 µg of pSP189 DNA in a transformation reaction [6]. MF was defined as the ratio of total mutants to total transformants [6].
Statistical analysis
Data are presented as mean ± SD. Statistical analysis was done using a Student’s t-test (SPSS for Windows, 12.0, SPSS Inc. Chicago, IL, USA). Differences were considered significant at p <0.05.
Results
Cytotoxicity and mutagenicity of NO• by reactor system
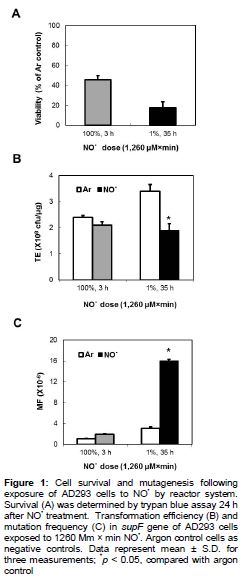
AD293 cells were exposed to NO• delivered by diffusion at a steady state concentration through gas-permeable tubing into the medium. When AD293 cells were exposed to 100 % NO• for 3 h or 1 % NO• for 35 h, cell viability were 46 and 18 %, resulting in cumulative 1260 μM × min total NO•, respectively (A).
Exposure to 100% NO• for 3 h or 1% NO• for 35 h also caused increases in supF MF compared to the Ar control, while corresponding decreases in TE were observed (B and C). When plasmids were treated with 100 % NO• for 3 h, the MF (1.9 ´ 10-6) was 1.9-fold higher than that of the Ar control (1 ´ 10-6). However, exposure to 1 % NO• for 35 h increased the MF 5.2-fold (16 ´ 10-6, p < 0.05), as compared with Ar control MF (3.1 ´ 10-6) ().
Viability, NO• production and mutagenesis of NO• by co-culture system
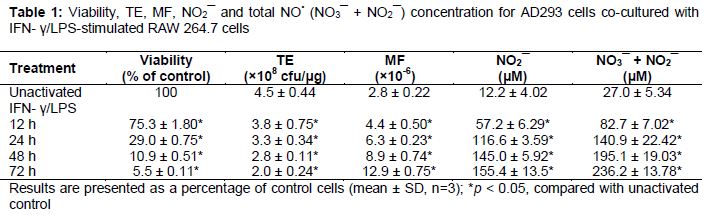
We determined the relative survival, concentration of NO• and mutation in the supF gene of AD293 cells co-cultivated with macrophages for 12, 24, 48 and 72 h (). Treatment of macrophages with IFN-γ and LPS for 24 h resulted in a statistically significant decreased AD293 cell viability (29 %), and 10.5-fold higher total NO• production compared with untreated control (p < 0.05) (). The MF (6.3 × 10-6) induced for 24 h was 2.3-fold greater (p < 0.05) than the spontaneous MFs (2.8 × 10-6) ().
NO• production, DNA damage and mutagenesis induced by SNP
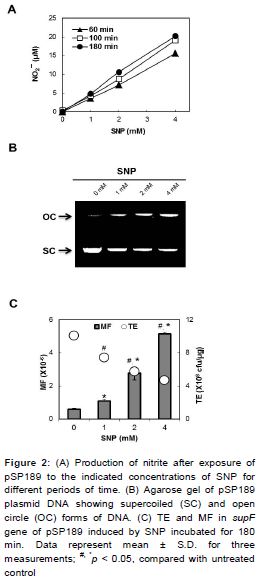
An increase in NO2¯ concentration in the medium was observed after exposure to a single concentration of SNP in a time-dependent manner (A). A dose dependent increase in NO2¯ concentration was also observed within the dose range of 1, 2 and 4 mM of SNP (A).
B shows results obtained from agarose gel electrophoresis of pSP189 plasmid after SNP exposure. For SNP, an increase in applied activity enhanced the plasmid damage, resulting in an increase in the open circular fractions and a decrease in the supercoiled fraction (B).
The MF of supF gene of pSP189 plasmid statistically increased following treatment with SNP, compared to that of the control (p < 0.05) (C). The MF induced by the treatment of 4 mM SNP was 8.8-fold higher than the spontaneous MF (p < 0.05) (C). Consistent with these results, the relative TEs statistically decreased in a dose-dependent manner after treatment with SNP (p < 0.05) (C).
Discussion
NO• is a short-lived molecule required for many physiological functions but high concentrations of NO• and ROS could lead to the DNA damage and potential genotoxic effects on hosts as well as neighboring cells. Several in vitro and in vivo model systems have been employed to examine the genotoxicity of NO• and ROS. Of these, a major mutation reporter system gene used in mammalian cells is the supF suppressor tRNA gene. Shuttle vector pSP189 has been used to determine mutations resulting from in vitro exposure to NO• and ROS followed by plasmid replication in bacterial and human cells, in order to determine possible effects of differences in repair and replication in the two host cells [6,8,12-21]. pSP189 contained several important features which make it useful for these studies. First, pSP189 carried a ‘signature sequence’ which distinguishes siblings and independent mutations in a shuttle vector [22]. Second, mutants in pSP189 represent several morphologically abnormal phenotypes. Third, pSP189 are useful for examining mutagenesis in various mammalian cell lines with DNA repair deficiencies. In vitro exposure of the plasmid to NO• gas resulted in increased mutation frequency, which was 15- and 44-fold higher than the spontaneous in E. coli MBM7070 and human AD293 cells, respectively [14].
The main purpose of the present study was to determine the effects of dose and dosing rate of NO• exposure on the levels of cytotoxicity, DNA damage and the frequency of mutations in the supF shuttle vector model as surrogates for in vivo studies of genotoxicity resulting from chronic inflammation. We have assessed the effects of NO• dose and dosing rate on the DNA damage and mutations induced in the supF gene by three different dosage mechanisms, enabling detection of mutations induced by diffusible agents: by stirred chamber in which preformed NO• diffused into the culture media from gas permeable tubing; by co-culture system in which target cells were physically separated from NO•-generating macrophages; and by exposure to SNP, which generates NO• spontaneously during decom-position.
When exposed to preformed 100 % NO• for 3 h, a cumulative dose of 1260 μM x min reduced the viability of AD293 cells at 24 h to 46 % and increased MF in the supF gene to 1.97 × 10−6 (p < 0.05), 1.9-fold statistically higher than background (). A comparable response in the treatment for 35 h with 1 % NO• at a same total dose (1260 μM × min) caused a statistically significant reduction in viability (18 %) and 5.3-fold higher MF, as compared with the argon control (), indicating NO• introduced slowly over time was more capable of inducing cytotoxicity and mutagenesis. Consistent with this interpretation, TE (an indication of total DNA damage) of plasmids exposed to 1% NO• for 35 h was lower than those exposed by 100 % NO• for 3 h. We also assessed the mutagenesis of the supF gene of pSP189 replicating in AD293 cells co-cultivated with activated macrophages and evaluated the relative importance of NO• as mediators of the genotoxicity induced.
Recent results show that NO• treatment reduced the percentage of viable cells with a time-dependent increase in NO• production and MF occurred in the supF gene of pSP189 replicating in AD293 cells co-cultivated with activated macrophages. An approximate 5-fold increase in MF of the supF gene in co-culture is similar to that observed in the experiments with preformed 1 % NO• for 35 h in reactor system (p < 0.05) (). SNP is an NO•-releasing compound that has been used as an anti-hypertensive agent since the 1920s [12]. The action of exogenous NO• donors can be similar to that observed in the case of macrophages, which paradoxically cause both growth and death of tumor cells [34]. As shown in , SNP has been shown to cause DNA strand breaks, and to cause a dose-dependent increase of the MF the supF locus in AD293 cells.
In work reported here, we found that both dose and dosing rate at which the supF gene is exposed to NO• strongly influence the DNA damage and the mutagenic potency in the gene. The systems used to introduce NO• in these experiments were designed to approximate conditions of exposure more physiologically relevant to chronic inflammation than bolus NO• additions, since cells in vivo are likely to be exposed over longer periods of time rather than to high concentrations for short periods. Hence, our findings provide important clues that dose and dosing rate NO• introduction may contribute to potential genotoxicity resulting from NO• formation in vivo.
Conclusion
The findings of this work indicate that NO• rate of dosing treatment affects genotoxic responses and these parameters may be of significance with regard to the potential genotoxicity resulting from NO• formation in vivo. Further studies will be required to elucidate precise underlying mechanisms of these effects and their potential relevance to NO•-induced genotoxicity in vivo.
Declarations
Acknowledgement
References
Archives


News Updates